| TOP>活動記録>講演会>第355回 | 一覧 | 次回 | 前回 | 戻る |
 |
|
|
|
|
第355回 邪馬台国の会(2017.1.22 開催)
| ||||
1.日本人の起源とフィリピン先住民の研究(尾本恵市先生)
|
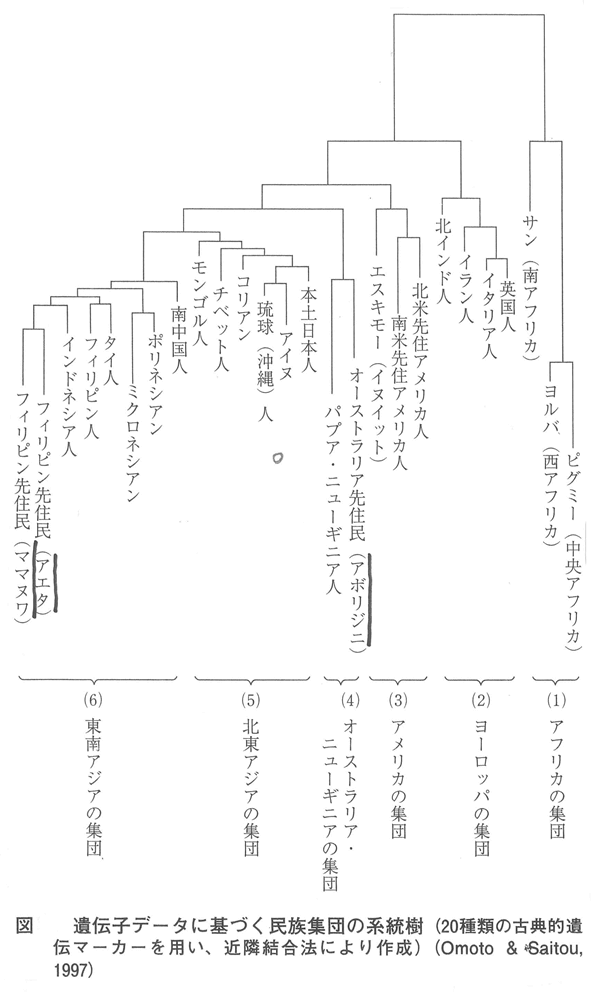
■日本人起源論の歴史 ■日本列島人(アイヌ、本土日本人、琉球人)の起源 しかし、遺伝子分析と合わなくなる。 タンパク質を使う古典的遺伝子マーカーによる民族集団の系統樹から、本土日本人と最初に分かれる民族がアイヌと琉球人である。その次がコリアンとなる。 ■縄文人と弥生人 しかしそのようにきちんと分けることができるのだろうか? ■『魏志倭人伝』に出てくる「侏儒国(しゅじゅこく)」とは? ■フィリピンと日本:自然と歴史とヒト ■「ネグリト」の起源(Small is beautiful) 最終氷河期(2~7万年前)の西太平洋にスンダランドがあった、そこはジャングルだったので、そこで、進化した人は小さくなった。小さいことはいいことだと言える。 (まだ、いろいろな講演内容がありましたが、おおまかな一部の話のみとしました。
|

2.倭人の弓、人類学と言語学(安本美典先生)
|
2.1 倭人の弓
また、福島大学教授の、工藤雅樹氏の著書『古代の蝦夷』(河出書房新社、1992年刊)にクマをねらう弓の文様の土器に、矢をつがえた弓と、クマのような動物を表現したもがのっている。青森県八戸市(韮窪遺跡)という場所からいって、アイヌの祖先にあたる人たちの用いたものであろう。
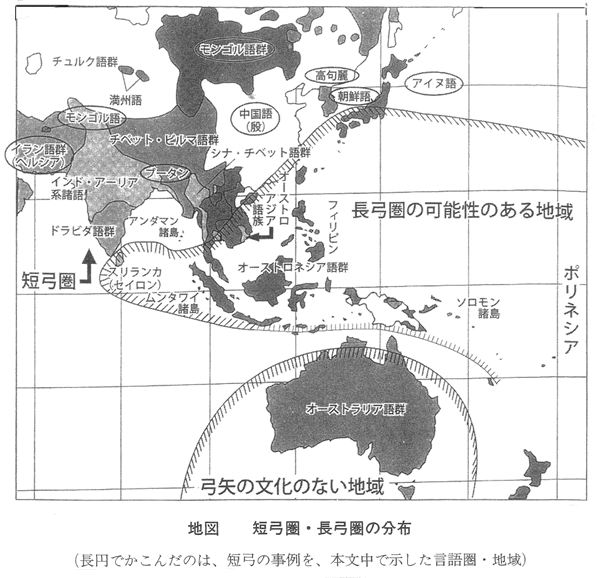
そして、長弓圏の可能性がある地域は下記のようだとして、以前に話をした。
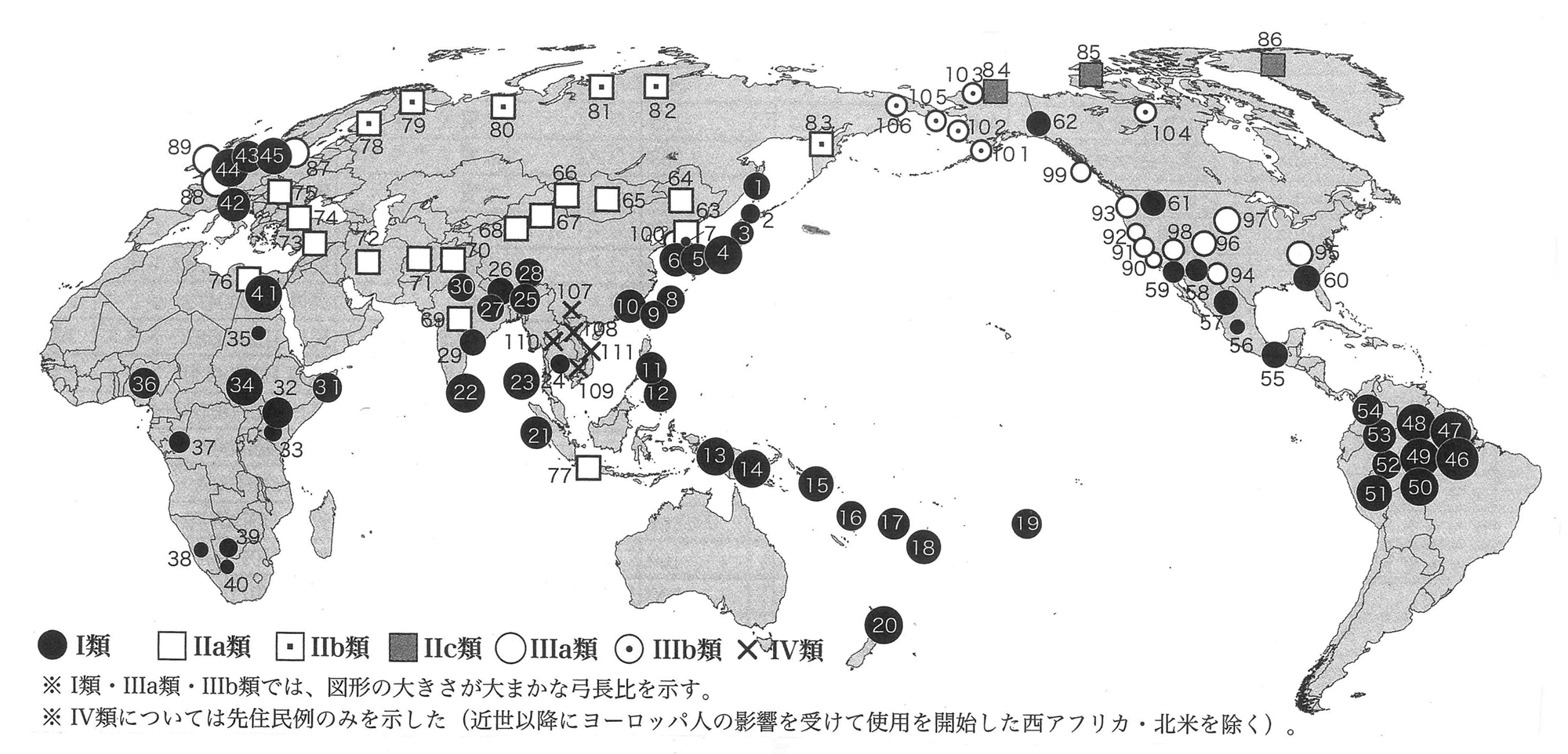
ここで、最近の話として、岡安光彦氏の和弓についての論文がある。 Ⅰ類のなかでも、ヒトの背丈を越える極端に長大な弓、Ⅰ類長大弓の分布は、オーストロネシア語族の分布域ないしその影響ときわめてよく重なる。太平洋の無人の島々に弓を伝えられたのは彼ら以外に存在しえない。Ⅰ類長大弓の伝播と拡散に、オーストロネシア語族が強く関与していたことは明白である。 拡散前のオーストロネシア語族の源郷(urheimat:Heine-Geldern1932)は、中国南部とみられる(J.ダイアモンド2000) 本論では、環太平洋地域における長弓の伝播がオーストロネシア語族と深い関係にあることを明らかにしたが、彼らの大規模な拡散の起点となった台湾は、職場の前任者『蘭嶼(らんしょ)とヤミと考古学』の筆者である故澤容一氏が、フィールドとして愛してやまなかった土地である。 太平洋およびインド洋周辺地域の長大なⅠ類弓は、海洋民(Sea-Nomad:Satherl995)たるオーストロネシア語族の大規模な拡散にともなって普及した。
太平洋西岸と南方の島嶼群に並んで、極端に長大なⅠ類弓が集中するのが、南アメリカ大陸のアマゾン川上流域一帯である。国立民族学博物館の標本のなかには、ブラジルのシングー川上流で採取された、228cmに及ぶ長弓がある。この地域では、ブラジルやベネズエラのヤノアマ(ヤノマミ)族、カマユラ族、ボドクト族、ペルーのトリオ族などが、長大なⅠ類弓を用いている。カマユラ族は漁労に利用する。ボドクト族はミトコンドリアDNA分析の結果、ポリネシア人と共通の遺伝子が発見されたが(Perikins2013)、ヨーロッパ人が釣り針をもたらすまでその存在を知らず、長大な弓と三叉鏃の矢で漁労をしていた(Metraux1946)。 オーストロネシア語族の拡散をめぐっては、言語統計学、人類学、考古学などの成果から、以下の仮説が有力視されている。紀元前3500年頃、台湾を起点に拡散が開始された。今日のアミ族、タイヤル族などオーストロネシア語族の言語を話す台湾先住民族がその子孫である。起元前3300年頃にはフィリピンへの入植が行われた。ここで、オーストロネシア語族は二手に分岐し、一群は西に向かい、インドネシアなど東南アジア島嶼群を席巻したのちに、マダガスカル島へ渡った。もう一群は東に向かい、紀元前1600年頃にオセアニア、前1200年頃にポリネシア、西暦500年頃にハワイ、西暦1000年頃にはニュー・ジーランドまで拡散した。サツマイモを、その栽培方法とともにポリネシアに持ち帰った。こうして「ポリネシア急行」と比喩される、急速かつ広範囲な拡散が完了した(P.ベルウッド1989、Bellwood1995、Gray2002)。
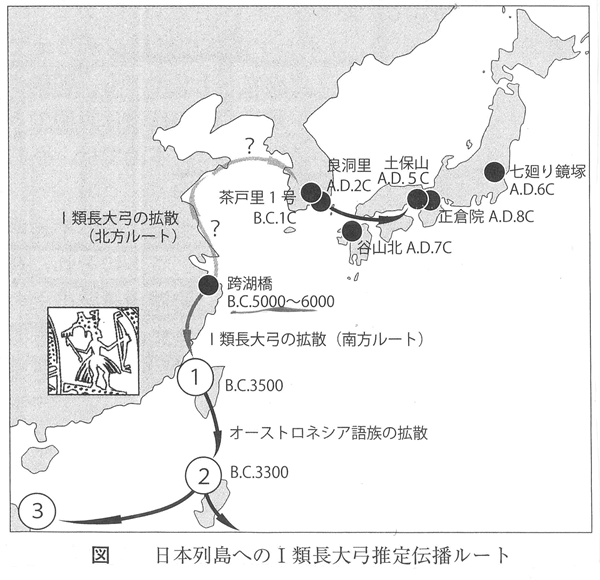
国立民族博物館には、先述した全長228cmに達する世界最長級の弓をはじめ、アマゾン川上流周辺で採集された先住民の長大な弓が多数収蔵されている。それらの弓の特徴は、太平洋型I類長大弓と酷似している。加えて、三叉鏃を付した長い漁労用の矢も相同である。南米大陸の弓と矢は、オーストロネシア語族がもたらした可能性がきわめて高い。 フィリピンやインドネシア海域の島嶼群の場合はどうか。この地域にはオーストロネシア語族の拡散以前に、すでにオーストラロイド系の狩猟採集民が先住していたが、太平洋型のⅠ類長大弓が広まった。オーストロネシア語族が先住民を人口的・文化的に淘汰してしまったからである(J.ダイアモンド2000)。フィリピンの狩猟漁労民ネグリートは、生き残ったオーストラロイド系の少数民であるが、言語的にはオースロネシア語族に同化され(P.ベルウッド1989)、生業もまたその影響を強く受けて、アウトリガー・カヌーを操り長大な弓と矢で漁労を営んでいる。 この地域では、180~190cm台あるいはそれ以上の長さに達するような、長大なⅠ類弓が、主体である。フィリピンのネグリート族は、原始和弓と同じ弓幹に樋と巻を施した長弓と、三叉(さんさ)鏃を付した長大な矢で、漁労に使用する(Krieger1926) 日本列島では縄文時代の短弓、弥生時代の比較的長い弓、長大な原始和弓など、さまざまな丸木弓が用いられてきた。朝鮮半島南部では、日本の弥生時代に相当する原三国時代(げんさんごくじだい)に、原始和弓に匹敵する長大なⅠ類弓が確認されている。起源前1世紀中頃の慶尚南道(けいしょうなんどう)昌原市(しょうげんし)茶戸里(ちゃどり)2号木棺墓(国立中央博物館2012)、2世紀前半頃の金海良洞里(きんかいりょうどうり)21号木槨墓(国立金海博物館2012)出土例である。 南下して東シナ海に目を転じると、杭州湾を臨む浙江省跨湖橋(ここきょう)遺跡から、新石器時代(紀元前5000~6000年頃)のⅠ類漆塗り丸木弓が出土している( 浙扛省文物考古研究所2004)。図面上の上側は折れて欠損し、下側もかなりの部分が剥離した状態で、残存長は121cmだが、一般的な弓のプロポーションから復元すると180cm前後の長大な弓であった可能性がある。詳しくは後述するが、跨湖橋(ここきょう)遺跡は稲作開始期の集落で、家畜化初期段階のブタ、丸木舟などが共伴している。 先に記した紀元前1世紀中頃の慶尚南道昌原市茶戸里1号木棺墓、紀元後2世紀前半頃の金海良洞里21号木槨墓という、いずれも朝鮮半島南部から出土したⅠ類の長大な漆弓を見ると、東シナ海沿岸の、跨湖橋遺跡から出土した、同じくⅠ類の漆弓との関連性に注目せざるをえない。紀元前後に朝鮮半島南部に出現し、遅れて日本列島に登場するⅠ類長大弓は、鳥霊思想にともなう他の文物などとともに、南方からもたらされた文化複合の構成要素ではないだろうか。 Ⅰ類長大弓の出現は、弥生文化が成立した後に起きた、時期的に異なる事象だからである。弥生文化の成立は、紀元前6世紀前後だろうが(田中2014)、I類長大弓が朝鮮半島南部に出現したのは紀元前1世紀頃以降、日本列島に出現したのは早くても弥生時代中期、確実には古墳時代に入ってからでのことである。
長弓の出発点は揚子江の河口の跨湖橋付近で、南は台湾へ行く、北は山東半島から朝鮮半島南岸から北九州へ伝わったとする。しかし南アメリカへ渡ったというところはどうだろうかといったところである。
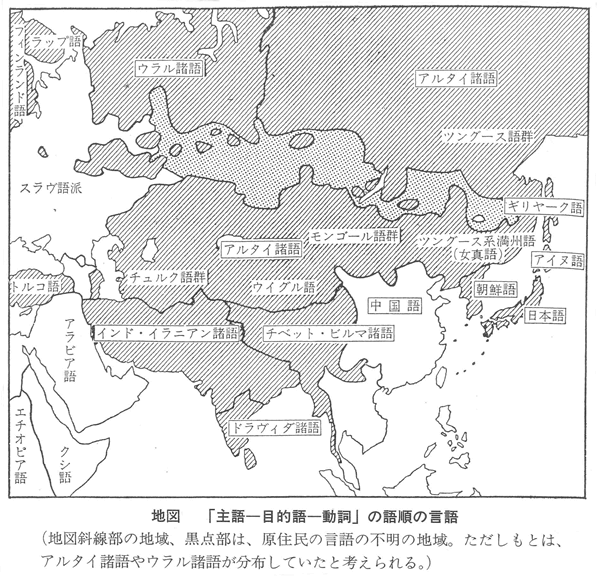
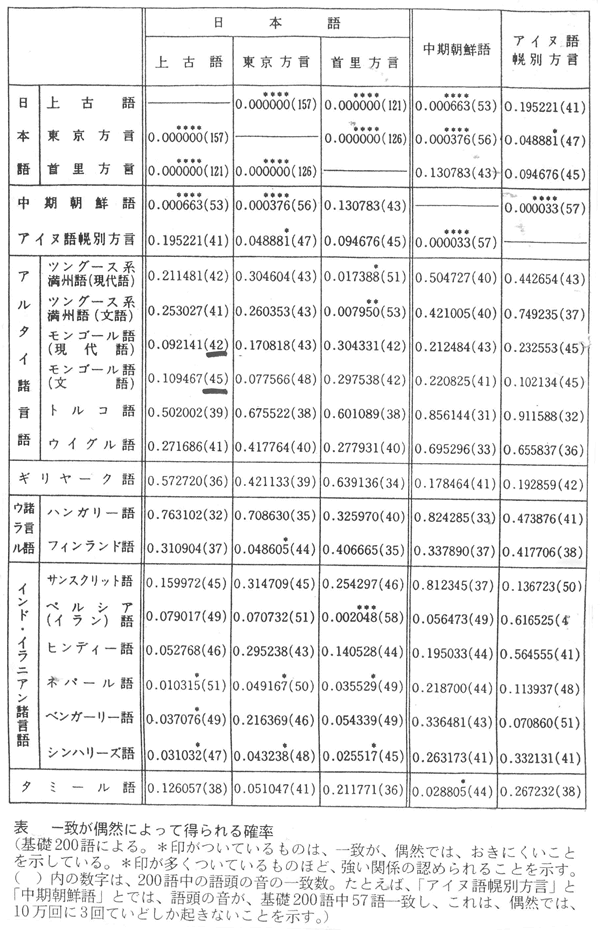
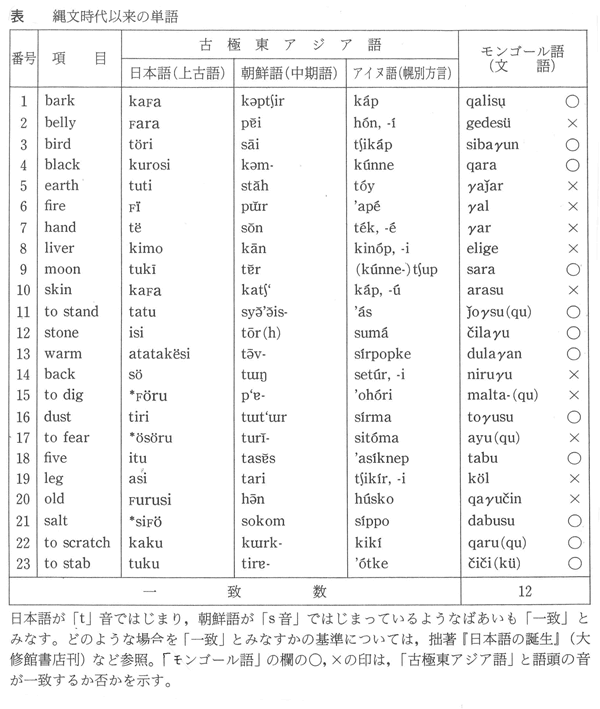
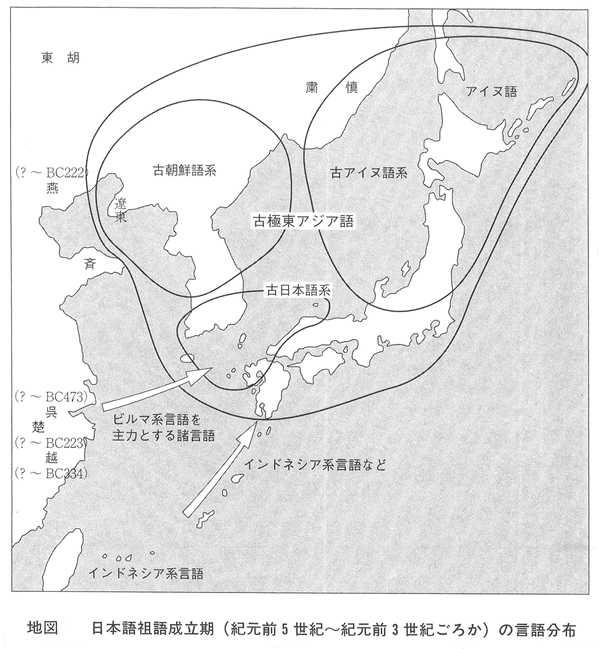
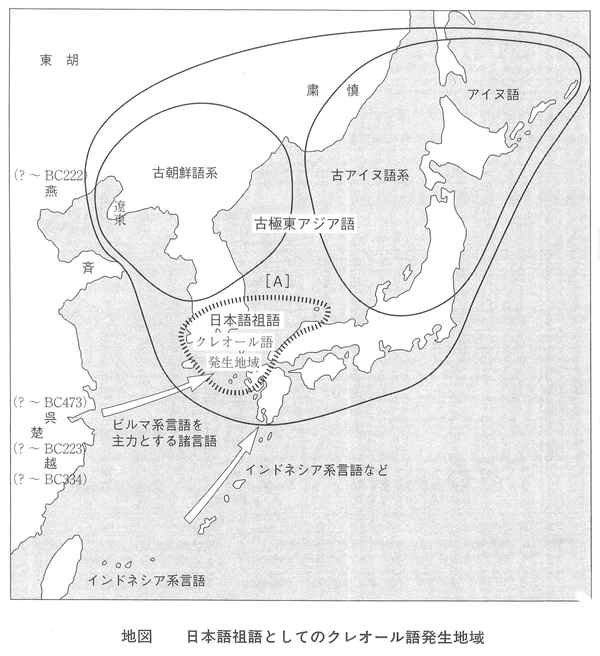
埴原和郎氏の「二重構造説」(尾本恵市氏による骨子) 尾本恵市氏の見解 ■語順 「語順の特徴に関するかぎり、日本語はユーラシアの中核的な『主語-目的語-動詞』圏の一環をなし、この地域との結びつきは疑うべくもない。また日本語の『目的語-動詞』的配列は、支配の方向性が驚くほど首尾一貫したもっともプロトタイプ(原型)に近い型といってもよい。また、この語順の型は、現存資料で溯りうる最古の時代から、基本的にはほとんど変化がなく、多くの過渡地域に見られる動揺や混合特徴は、漢語やヨーロッパ語の影響による一部の現象を除いて、まったく認められない。日本語はこの点で、インド亜大陸のドラヴィダ諸語、アジア北・東部のアルタイ諸語と共に、『目的語-動詞』言語圏の中のもっとも安定したタイプに属し、他方、東南アジアとオセアニア(オーストラリア、ニューギュア、メラネシア、ポリネシア、ミクロネシア)を中心とする『動詞-目的語』言語圏とは明確な一線を画している。」 ■アイヌ語と朝鮮語との関係 日本上古語、中期朝鮮語、アイヌ語紋別方言はモンゴル語と偶然以上の一致はないが、日本上古語、中期朝鮮語、アイヌ語紋別方言に共通する単語23をモンゴル語と比較すると、一致するものが多くなる。つまり古い単語だけとれば、一致する確率が上がる。 ラテン語はルーマニア語、イタリア語、フランス語などに分かれたが、日本語は古極東アジア語などをベースにインドネシア系言語、クメール系言語、ビルマ、中国語などが入ってきて成立した。 この日本語の成立について、地図上に示すと下図の「日本祖語成立期(紀元前5世紀~紀元前3世紀ごとか)の言語分布」のようになる。 これは、日本祖語としてのクレオール語となったと考えられる。「日本祖語としてのクレオール語発生地域」参照
上古日本語と偶然以上の一致がみられるものの3番目に、インドネシア語がある。
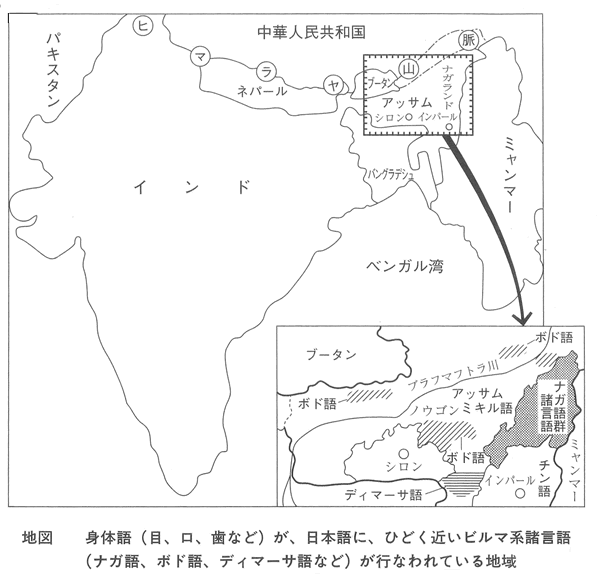
ポド語とか、ナガ語が身体語に関して、上古日本語に近い。
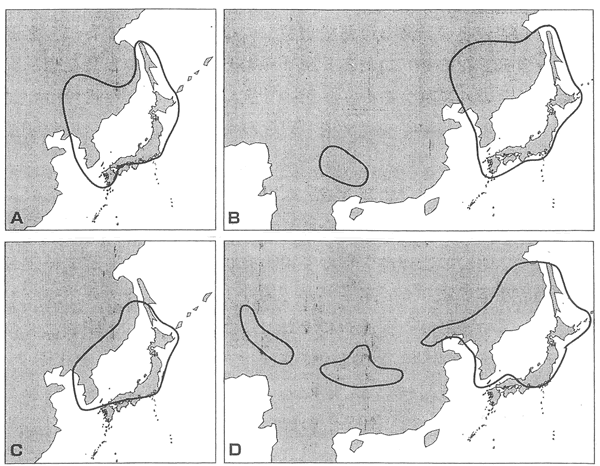
このポド語とか、ナガ語を話す人達はインドのアッサム地方の住民である。 生物の分布から、面白い現象がある。 C、D陸上高等植物の場合
このように、日本海を取り囲むように分布しているものが、ヒマラヤの方にも存在する。
|








3.対談
|
■尾本先生 その他いろいろな話がありましたが、省略します。 |

| TOP>活動記録>講演会>第355回 | 一覧 | 上へ | 次回 | 前回 | 戻る |